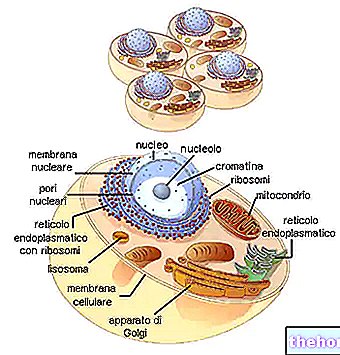
يبدأ تركيب الأحماض الدهنية من أنزيم الأسيتيل المساعد A ويتوافق تقريبًا مع المسار العكسي لتحللها ؛ في تخليق الأحماض الدهنية ، تتم إضافة سلسلة من شظايا البيكربونات إلى الإنزيم المساعد أسيتيل البادئ.
إن تخليق الأحماض الدهنية هو سيتوبلازمي بالكامل (أي الإنزيمات التي تحفز هذا التركيب موجودة في السيتوبلازم). إنزيم الأسيتيل A المستخدم في السيتوبلازم لتخليق الأحماض الدهنية هو من أصل ميتوكوندريا: يتم نقل جزء صغير من خلال الكارنيتين ، عن طريق عمل اثنين من إنزيمات ترانسفيراز الأسيل (واحد سيتوبلازم والآخر ميتوكوندريا) وإنزيم ترانسيلوكاز. جزء من الأسيتيل يتم الحصول على الإنزيم المساعد A من أصل الميتوكوندريا من خلال طريق متخصص: سترات لياز (الاسم مشتق من الإنزيم الأول لهذا المسار).
مشتق أسيتيل أنزيم أ الموجود في الميتوكوندريا من تحلل السكر بعد عمل بيروفات ديهيدروجينيز ؛ يخضع الإنزيم المساعد أسيتيل أ لتأثير إنزيم سيترات سينسيز: يحفز هذا الإنزيم تكوين السترات عن طريق تفاعل أسيتيل الإنزيم المساعد أ مع أوكسالأسيتات. إذا كانت دورة كريبس قادرة على تلبية احتياجات الطاقة ، فإنها تبدأ من السترات (الكمية غير الضرورية في دورة كريبس) يمكن أن تترك الميتوكوندريا وتصل إلى السيتوبلازم ، حيث إنزيم لياز السترات ، الذي ينفق الطاقة ، يحولها مرة أخرى إلى أنزيم أسيتيل أ وأوكسلو أسيتات. وبهذه الطريقة يمكن أن يكون هناك أنزيم أسيتيل أ متاح في السيتوبلازم ؛ ومع ذلك ، يجب إعادة أوكسالو أسيتات التي يتم تكوينها إلى الميتوكوندريا من أجل أن تكون متاحة مرة أخرى لإنزيم سينثيز السترات.
ثم يتم تحويل أوكسالو أسيتات إلى مالات بفعل الإنزيم نازعة هيدروجين مالات السيتوبلازميك (يتم إنفاق NADH السيتوبلازمي): المالات هو مستقلب نافذ ويمكنه إعادة دخول الميتوكوندريا حيث ، تحت تأثير إنزيم نازعة الهيدروجين في الميتوكوندريا ، يتم إعادة تحويله إلى oxaloacetate (يتم الحصول على NADH أيضًا) ؛ يمكن للمريض السيتوبلازمي ، بدلاً من ذلك ، الخضوع لعمل إنزيم الماليك ، الذي ينفذ نزع الكربوكسيل ونزع الهيدروجين ، ليتم تحويله إلى بيروفات. يعمل إنزيم الماليك على NADP + (وهو مشابه لنيكوتيناميد أدينينوكليوتيد ولكن ، على عكس هذا ، فإنه يحتوي على مجموعة فسفورية في مجموعة الهيدروكسيل الثانية على إحدى وحدتي الريبوز) لذلك في الممر من مالات إلى البيروفات ، يتم إنتاج NADPH ( الذي يستخدم في التخليق الحيوي) ثم يدخل البيروفات إلى الميتوكوندريا حيث يتحول إلى أوكسال أسيتات عن طريق عمل بيروفات كاربوكسيلاز أو أسيتيل أنزيم أ من خلال بيروفات ديهيدروجينيز.
دعونا نرى مثالاً: هناك حاجة لثمانية جزيئات من الإنزيم المساعد أسيتيل لتخليق حمض البالمتيك (سلسلة تحتوي على ستة عشر ذرة كربون) ولكن يتم استخدام واحدة منها فقط على هذا النحو: يتم تحويل سبعة جزيئات من الإنزيم المساعد أسيتيل إلى أنزيم مالونيل أ عن طريق "الإنزيم" أسيتيل أنزيم A كربوكسيلاز (يستخدم هذا الإنزيم جزيء ثاني أكسيد الكربون وله البيوتين كعامل مساعد).
يمكن أن يوجد إنزيم الأسيتيل الإنزيم A carboxylase في شكل مشتت غير نشط تقريبًا وشكل تجميع نشط (حوالي عشرين وحدة) ؛ يحدث الانتقال من الشكل المشتت إلى الشكل الكلي عندما يوجد في السيتوبلازم "تركيز عالٍ من السيترات: السيترات هو معدل إيجابي لأنزيم أسيتيل إنزيم الكربوكسيلاز.
يحتوي إنزيم الأسيتيل A carboxylase على مُعدِّلات أخرى إيجابية (الأنسولين) وسلبية (الجلوكاجون والأدرينالين وأنزيم أسيل الإنزيم أ).
سنقوم بتحليل تخليق الأحماض الدهنية في بكتيريا الإشريكية القولونية التي يحدث فيها هذا التوليف بفعل سبعة بروتينات متميزة ؛ في الخلايا حقيقية النواة ، تتشابه الآلية التي يتم من خلالها تخليق الأحماض الدهنية مع تلك الخاصة بالبكتيريا ، ولكن في حقيقيات النوى ، يتم تجميع الإنزيمات السبعة المسؤولة عن التوليف في مجمعين متعددي الإنزيمات A و B.
في البكتيريا ، ترمز سبعة جينات متميزة لـ:
- ACP (بروتين ناقل الأسيل) ؛
- ACP-acetyl transacetylase ؛
- ACP.مالونيل ترانس أسيتيلاز ؛
- β-keto-acyl-ACP synthase (إنزيم التكثيف) ؛
- اختزال β-keto-acyl-ACP ؛
- D-β-hydroxy-acyl dehydratase ؛
- enoil-ACP منقح.
في حقيقيات النوى ، هناك رمزان جينيان لـ:
الوحدة الفرعية أ
ACP.
انزيم التكثيف
اختزال β-keto-acyl-ACP.
الوحدة الفرعية ب
ACP-acetyl transacetylase ؛
ACP-malonyl transacetylase ؛
D-β-hydroxy-acyl dehydratase ؛
enoil-ACP منقح.
يتم ترتيب البروتينات السبعة من Escherichia coli بحيث يوجد بروتين مركزي (ACP) وستة أخرى حوله.
تشترك مجموعتان من السلفهيدريل في تأثيره الأنزيمي: واحدة تنتمي إلى السيستين والأخرى تنتمي إلى الذراع الطويلة للفوسفوبانثيئين ؛ يرتبط ACP بالركيزة التي ، من خلال ذراع الفوسفوبانثيثين ، تتلامس مع الإنزيمات الأخرى التي تكون بالتالي قادرة على القيام بعملها الأنزيمي.
يرتبط Acetyl coenzyme A (عن طريق ACP acetyl transacylase) بإنزيم ACP (بشكل أكثر دقة إلى كبريت السيستين الذي يشكل مشتق السيستيل) ويتم إطلاق الإنزيم المساعد A ؛ ثم يتدخل ACP-malonyl Transacylase مما يحفز هجوم malonyl على phosphopanthetheine (أيضًا في هذه العملية يتم تحرير الإنزيم المساعد A الذي كان مرتبطًا في البداية بالمالونيل).
تتضمن الخطوة التالية سينسيز β-keto-acyl ACP وهو إنزيم مكثف: يسمح بالاندماج بين الهيكلين العظميين ؛ يتم إزالة الكربوكسيل بسهولة من malonyl ويتم تكوين كربونيل من مشتق الأسيتيل السيستين: يتم تحرير السيستين ويتم تكوين مشتق β-keto (acetyl acetyl) phosphopantethin.
بعد ذلك ، يتدخل إنزيم β-keto-acyl-ACP الذي يقلل الكربونيل بشكل أكبر إلى إنزيم ACP (يتكون الهيدروكسيد بواسطة NADPH الذي يتم تقليله إلى NADP +).
الآن ، يعمل 3-hydroxy-acyl ACP dehydratase (يحدث الجفاف) مما يؤدي إلى تكوين نظام غير مشبع (ألكين).
تتضمن العملية التالية enoyl-ACP-reductase (تقوم بعملية الهدرجة: يتم تشكيل الألكان ويتم تقليل NADPH إلى NADP +).
تتضمن المرحلة الأخيرة تحويل منتج الأسيل الذي تم الحصول عليه من الدورة الأولى إلى مركب قادر على بدء دورة ثانية: ينقل إنزيم ترانساسيلاز الأسيل إلى السيستين ، تاركًا موقع البانتيثين الذي سيكون الآن على استعداد لربط آخر. مالونيل.
في الأكسدة β ، يتم استخدام جزيء FAD للحصول على إنزيم α-المستقلب غير المشبع عبر الإنزيم A عن طريق نزع الهيدروجين ؛ في تركيب الأحماض الدهنية ، بدلاً من ذلك ، يتم استخدام جزيء NADPH لإحداث رد فعل معاكس.
عادة ، يتم تصنيع الأحماض الدهنية مع ستة عشر ذرة كربون ، ولكن يمكن أيضًا إنتاج الأحماض الدهنية مع ثمانية عشر أو عشرين أو اثنتين وعشرين ذرة كربون ؛ ثم يتم أسترة الأحماض الدهنية لتشكيل الدهون الثلاثية مع الجلسرين المنشط (أي الجلسرين 3 - الفوسفات). يمكن الحصول على الأخير من ثنائي هيدروكسي أسيتون فوسفات بفعل الإنزيم نازعة هيدروجين فوسفات الجلسرين أو من الجلسرين عبر الإنزيم كيناز الجلسرين.
يجب إرسال الأحماض الدهنية المركبة إلى الأنسجة الدهنية ؛ يتم نقلها في مجرى الدم على شكل دهون ثلاثية أو ، جزئيًا ، باستخدام بروتين ناقل وهو الألبومين.