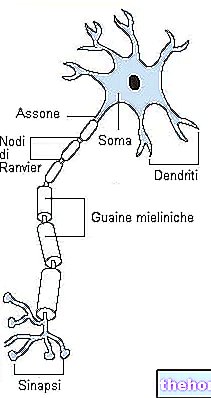
الجليكوجين هو جزيء ضخم (كتلة جزيئية تبلغ حوالي 400 مليون دالتون) من α-glucose حيث توجد روابط α-1،4 glycosidic وتشعبات بنسبة 1:10 ، بسبب روابط α-1،6 glycosidic.
يشكل الجليكوجين مادة احتياطية ويتحلل ويعاد تكوينه باستمرار ؛ في كتلة خلايا الجسم بالكامل ، يوجد حوالي 100 جرام من الجليكوجين: يوجد معظمه في الكبد حيث يكون متحركًا ، وبالتالي يمكن استخدامه كاحتياطي للأعضاء الأخرى (الجليكوجين في العضلات غير متحرك).
إن الإنزيمات التي تحفز تحلل الجليكوجين وتكوينه كلها موجودة في السيتوبلازم ، لذلك هناك حاجة إلى نظام تنظيم يجعل أحد المسارات غير نشط عندما يكون الآخر نشطًا: إذا كان هناك جلوكوز متاح ، يتم تحويل الأخير إلى جليكوجين (الابتنائية) وهو احتياطي ، بالعكس ، إذا كان c "مطلوبًا للجلوكوز ، فإن الجليكوجين يتحلل (تقويض).

الإنزيم المتورط بشكل رئيسي في تكسير الجليكوجين هو فسفوريلاز الجليكوجين؛ هذا الإنزيم قادر على شق رابطة جليكوسيدية α-1،4 باستخدام أورثوفوسفات غير عضوي كعامل حل: يحدث الانقسام بطريقة الفسفور ويتم الحصول على الجلوكوز 1-فوسفات.
عند خمس أو ست وحدات من نقطة التفرع ، لم يعد إنزيم الجليكوجين فسفوريلاز قادرًا على العمل ، وبالتالي ينفصل عن الجليكوجين ويتم استبداله بإنزيم مشتت وهو عبارة عن ناقل: في الموقع التحفيزي لهذا الإنزيم c يوجد "هيستيدين" الذي يسمح بنقل ثلاث وحدات من السكريد إلى أقرب سلسلة جليكوسيدية (يهاجم الهيستيدين الكربون الأول لجزيء الجلوكوز). والإنزيم المذكور للتو هو جليكوزيل ترانسفيراز؛ في نهاية عمل هذا الإنزيم ، تبقى وحدة جلوكوز واحدة فقط على السلسلة الجانبية حيث يرتبط أول كربون بالكربون السادس للجلوكوز في السلسلة الرئيسية. ويتم تحرير آخر وحدة جلوكوز في السلسلة الجانبية بفعل "إنزيم α-1،6 جليكوزيداز (يشكل هذا الإنزيم الجزء الثاني من إنزيم إزالة الصفاء) ؛ بالنظر إلى أن الفروع في الجليكوجين تكون بنسبة 1:10 ، من التحلل الكامل للجزيء الكبير نحصل على حوالي 90٪ من الجلوكوز 1-فوسفات وحوالي 10٪ من الجلوكوز.
يسمح عمل الإنزيمات المذكورة أعلاه بإزالة سلسلة جانبية من جزيء الجليكوجين ؛ يمكن تكرار نشاط هذه الإنزيمات حتى يحدث التدهور الكامل للسلسلة.
دعونا نفكر في خلية كبدية. الجلوكوز (يتم امتصاصه من خلال النظام الغذائي) ، عندما يدخل الخلية يتحول إلى جلوكوز 6 فوسفات وبالتالي يتم تنشيطه. الجلوكوز 6 فوسفات ، بفعل فسفوجلوكوموتاز، يتحول إلى جلوكوز 1- فوسفات: هذا الأخير هو مقدمة غير فورية للتخليق الحيوي ؛ في التخليق الحيوي ، يتم استخدام شكل منشط من السكريات والذي يمثله سكر مرتبط بثنائي فوسفات: عادة يوريدي فوسفات (UDP). الجلوكوز 1- الفوسفات هو ثم يتم تحويله إلى UDP- جلوكوز ، هذا المستقلب تحت تأثير سينثيز الجليكوجين وهو قادر على ربط UDP-glucose بنهاية غير مختصرة للجليكوجين المتنامي: يتم الحصول على الجليكوجين المطول لوحدة الجلوكوزيد و UDP. يتم تحويل UDP بواسطة إنزيم ثنائي فسفوكيناز النوكليوزيد إلى UTP الذي يعود إلى الدورة الدموية.
يحدث تحلل الجليكوجين بفعل فسفوريلاز الجليكوجين الذي يطلق جزيء الجلوكوز ويحوله إلى جلوكوز 1 فوسفات. بعد ذلك ، يقوم الفوسفوجلوكوموتاز بتحويل الجلوكوز 1-فوسفات إلى الجلوكوز 6-فوسفات.
يتم تصنيع الجليكوجين ، قبل كل شيء ، في الكبد والعضلات: يوجد في الكائن الحي 1-1.2 هيكتوجرام من الجليكوجين موزعة في جميع أنحاء كتلة العضلات.
يمثل الجليكوجين في الخلية العضلية احتياطيًا للطاقة لهذه الخلية فقط بينما يمثل الجليكوجين الموجود في الكبد أيضًا احتياطيًا للأنسجة الأخرى ، أي يمكن إرساله ، مثل الجلوكوز ، إلى خلايا أخرى.
يتم بعد ذلك إرسال الجلوكوز 6-فوسفات الذي يتم الحصول عليه في العضلات من تحلل الجليكوجين ، في حالة الحاجة إلى الطاقة ، إلى تحلل الجلوكوز ؛ في الكبد ، يتم تحويل الجلوكوز 6-فوسفات إلى جلوكوز بفعل جلوكوز 6 فوسفات فوسفاتيز (إنزيم مميز لخلايا الكبد) وينتقل إلى مجرى الدم.
يعمل كل من Glycogen synthase و glycogen phosphorylase على الوحدات غير المختزلة من الجليكوجين لذلك يجب أن تكون هناك إشارة هرمونية تطلب تنشيط أحد المسارات وحجب الآخر (أو العكس بالعكس).
في المختبر كان من الممكن إطالة سلسلة الجليكوجين عن طريق استغلال فوسفوريلاز الجليكوجين واستخدام الجلوكوز 1-فوسفات بتركيز عالٍ جدًا.
في الخلايا ، يحفز فوسفوريلاز الجليكوجين فقط تفاعل التحلل لأن تركيزات المستقلبات مثل تحويل توازن التفاعل التالي إلى اليمين (أي نحو تحلل الجليكوجين):

دعونا نرى آلية عمل فوسفوريلاز الجليكوجين: يرتبط أكسجين الأسيتال (الذي يعمل كجسر بين وحدات الجلوكوز) بهيدروجين الفوسفوريل: يتم تكوين وسيط التفاعل بواسطة carbocation (على الجلوكوز الذي هو كل شيء " الأطراف) التي يرتبط بها الفسفوريل (Pi) بسرعة كبيرة.
يتطلب فسفوريلاز الجليكوجين عاملاً مساعدًا وهو فوسفات البيريدوكسال (هذا الجزيء هو أيضًا عامل مساعد في الترانساميناسات): يحتوي على فسفوريل بروتوني جزئيًا (فوسفات البيريدوكسال محاط ببيئة كارهة للماء مما يبرر وجود البروتونات المرتبطة به). الفوسفوريل (Pi) قادر على نقل البروتون إلى الجليكوجين لأن هذا الفوسفوريل يستعيد البروتون من الفسفوريل البروتوني جزئيًا لفوسفات البيريدوكسال. احتمالية أن يفقد الفسفوريل بروتونه عند درجة الحموضة الفسيولوجية ويظل منزوعًا تمامًا من البروتون منخفض جدًا.
دعونا الآن نرى كيف يعمل الفوسفوجلوكوموتاز. يقدم هذا الإنزيم ، في الموقع التحفيزي ، بقايا من السيرين الفسفوري ؛ ينتج السيرين الفوسفوريل إلى الجلوكوز 1-الفوسفات (في الموضع السادس): يتكون الجلوكوز 1،6-بيسفوسفات لفترة قصيرة ، ثم يتم إعادة فسفرة السيرين مع أخذ الفوسفوريل في الموضع الأول. يمكن أن يعمل Phosphogluco mutase في كلا الاتجاهين ، أي تحويل الجلوكوز 1-فوسفات إلى جلوكوز 6-فوسفات أو العكس ؛ إذا تم إنتاج الجلوكوز 6-فوسفات ، فيمكن إرساله مباشرة إلى تحلل الجلوكوز ، في العضلات ، أو تحويله إلى جلوكوز في الكبد.
الانزيم uridyl phosphogluco transferase (أو UDP الجلوكوز بيروفوسفوريلاز) يحفز تفاعل نقل الجلوكوز 1-فوسفات إلى UTP عن طريق التعلق بالفوسفوريل أ.
الإنزيم الموصوف للتو هو بيروفوسفوريلاز: هذا الاسم يرجع إلى حقيقة أن رد الفعل المعاكس للإنزيم الموصوف للتو هو تفاعل بيروفوسفوريلاز.
الجلوكوز UDP ، الذي تم الحصول عليه كما هو موصوف ، قادر على إطالة سلسلة الجليكوجين ، بواسطة وحدة أحادية السكاريد.
من الممكن جعل التفاعل يتطور نحو تكوين جلوكوز UDP عن طريق التخلص من منتج هو بيروفوسفات ؛ الانزيم بيروفوسفاتيز يحول البيروفوسفات إلى جزيئين من أورثوفوسفات (التحلل المائي لأنهيدريد) وبذلك ، يحافظ على تركيز بيروفوسفات منخفضًا جدًا لجعل عملية تكوين جلوكوز UDP مفضلة ديناميكيًا حراريًا.
كما ذكرنا ، فإن الجلوكوز UDP ، بفضل عمل الجليكوجين سينثيز ، قادر على إطالة سلسلة الجليكوجين.
ترجع التداعيات (بنسبة 1:10) إلى حقيقة أنه عندما تتكون سلسلة الجليكوجين من 20-25 وحدة ، يتدخل إنزيم متفرع (له "هيستيدين في موقعه الحفاز) ، قادرًا على نقل سلسلة من 7-8 وحدات جليكوسيدية إضافية من 5-6 وحدات: وبالتالي يتم إنشاء تفريع جديد.
لأسباب تتعلق بالمنشأ العصبي أو إذا كانت الطاقة مطلوبة بسبب المجهود البدني ، يتم إفراز الأدرينالين من الغدد الكظرية.
الخلايا المستهدفة من الأدرينالين (والنورأدرينالين) هي خلايا الكبد والعضلات والأنسجة الدهنية (في الأخير هناك تحلل الدهون الثلاثية وتداول الأحماض الدهنية: وبالتالي ، يتم إنتاج الجلوكوز في الميتوكوندريا 6-فوسفات ، ليكون يتم إرساله إلى تحلل الجلوكوز ، بينما في الخلايا الدهنية ، يتم تحويل الجلوكوز 6 فوسفات إلى جلوكوز عن طريق عمل إنزيم الجلوكوز 6 فوسفات الفوسفات ويتم تصديره إلى الأنسجة).
دعونا نرى الآن طرق عمل الأدرينالين. يرتبط الأدرينالين بمستقبل يوضع على غشاء الخلية (الخلايا العضلية والخلايا الكبدية) وهذا يحدد ترجمة الإشارة من الخارج إلى داخل الخلية.يتم تنشيط بروتين كيناز الذي يعمل في نفس الوقت على الأنظمة التي تنظم تخليق وتدهور الجليكوجين:
يوجد سينسيز الجليكوجين في شكلين: شكل مزوَّد فسفرة (نشط) وشكل فسفوري (غير نشط) ؛ بروتين كيناز فسفوريلات سينسيز الجليكوجين ويمنع عمله.
يمكن أن يوجد فوسفوريلاز الجليكوجين في شكلين: شكل نشط يوجد فيه سيرين فسفري وشكل غير نشط يتم فيه نزع الفسفرة من السيرين. يمكن تنشيط فوسفوريلاز الجليكوجين بواسطة الإنزيم الجليكوجين فسفوريلاز كيناز. يكون الجليكوجين فسفوريلاز كيناز نشطًا إذا كان مُفسفرًا وغير نشط إذا تم نزع الفسفرة منه ؛ يحتوي بروتين كينيز كركيزة الجليكوجين فسفوريلاز كيناز ، أي أنه قادر على الفسفرة (وبالتالي تنشيط) الأخير الذي بدوره ينشط فوسفوريلاز الجليكوجين.
بمجرد انتهاء إشارة الأدرينالين ، يجب أن ينتهي تأثيرها على الخلية أيضًا: تتدخل إنزيمات الفوسفاتيز في أنواع البروتين.